Body Defence against Ringworm (Tinea)
Introduction
Ringworm, or tinea, is a fungal disease that affects the epidermal and dermal layers of the skin of humans and animals. ‘Ringworm’ is an historic term which was given by early scientists as they considered the causative agents to be worms and because of the characteristic ring-shaped lesions formed on affected skin or scalps.
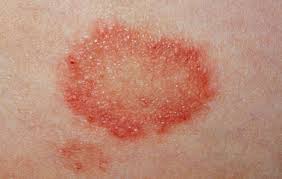
Source: http://www.nhs.uk/conditions/Ringworm/Pages/Introduction.aspx
Fungi are eukaryotic, free-living or parasitic micro-organisms that mostly require a weakness in the body’s immune system to establish a disease; thus virtually every fungal infection is opportunistic. In humans, it is mostly young children who are infected. Fungal infections are now growing at a rapid rate which is probably due to an increase in the number of immune-compromised patients (Detandt and Nolard, 1995).
The tinea fungus is transmitted by direct person-to-person contact, by sharing items such as combs, brushes, hats and pillows with infected persons or by touching contaminated surfaces (Piιrard et al., 1996; Bronson et al., 1983; Hay et al., 1996). Cases of infected people have been reported all over the world but are more prevalent in areas where there are poor hygienic standards. Dermatophytes, a group of closely-related fungi, are divided into three genera: Trichophyton, Microsporum and Epidermophyton. These are the causative agents of ringworm. They invade the inner tissues of the skin and utilise keratin which stimulates the body’s defence system to start an immune response to get rid of invaders. The immune system is responsible for defending the body against foreign invaders and functions by means of two mechanisms:
1. Non-specific immunity:
This is the first line of defence and is provided by mechanisms that form an immediate natural barrier to infections like tinea. Local defence mechanisms such as unsaturated transferrin, phagocytosis by polymorphonuclear leucocytes and medium chain-length fatty acids which are produced in sebum are inhibitory to dermatophyte growth (Hay, 1997).
2. Specific immunity:
This is mediated through two arms: humoral by means of B-lymphocytes and cellular by means of T-lymphocytes.
a. Humoral immunity:
Circulating antibodies have been demonstrated in the sera of humans and animals with natural or experimental ringworm infection (Grappel et al., l974). However, there is no uniformity of opinion as to the protective role of antibodies in dermatophytosis. Many investigators have failed to correlate antibody titre with the clinical picture or the development of immunity. This discrepancy may be due in part to different experimental designs and various ways of preparing test antigens and the time of collection of the sera. To date, evidence that antibodies have any protective role is largely inconclusive.
b. Cell-mediated immunity:
Immunity to intracellular facultative parasites is conferred by T-lymphocytes. T-cells differentiate in the thymus and serve a number of functions. These include helping B-cells to make antibodies, killing virally-infected cells, regulating the level of the immune response and stimulating activity of other effector cells like macrophages to engulf and kill micro-organisms. T-cells are divided into three sub-sets with two important sub-populations: T-helper and T-cytotoxic/ T-suppressor.
Stimulated T-cells proliferate and differentiate to give rise to two major effector mechanisms: generation of cytotoxic cells and release of lymphokines. The lymphokines are biologically active factors released by lymphocytes that may affect target cells directly or indirectly by modulating the behaviour of other effector cells (Delves et al., 2011). Immunological priming of a host and subsequent contact with the sensitising antigen boosts the secondary immune response, but the reaction may be severe and lead to tissue damage. This state is described as hypersensitivity. Immediate and delayed-type hypersensitivity will be briefly described here.
In immediate hypersensitivity, the antigen reacts with the IgE antibody that is bound to mast cells. This leads to the degranulation of mast cells and the release of histamine and other mediators of anaphylaxis. In delayed-type hypersensitivity, the antigen stimulates T-cells to release lymphokines which regulate a series of cellular reactions necessary to eliminate the antigen. The reaction is characterised by redness and induration at the site of antigen inoculation and appears after several hours (Delves et al., 2011).
Cell-mediated immune responses are considered to play a major role in fighting tinea (Grappel, l981). The development of cell-mediated immunity during experimental murine dermatophytosis has been studied. Calderon and Hay (l984 a,b) induced acute and chronic infection in BALB/c mice with Tricophyton quinckeanum and investigated immune responses by lymphocyte transformation and cell transfer experiments. Mice were irradiated to kill the lymphocytes and then T-helper cells from both acutely or chronically infected mice were transferred to them.
These experiments demonstrated that T-cells play an important role in the specific response to a fungal antigen and also to the suppressor activity. Further, it was found that adoptive immunity was abrogated by serum from chronically infected donors. The serum factor was considered to be a circulating antigen and in further studies a specific dermatophyte antigen alone eliminated the adoptive transfer. Antigens derived from T- species containing certain reactive epitopes (probably glycopeptides) interfered with T-cell-mediated immunity.
The Skin Test:
Resistance to reinfection by a dermatophyte correlates well with the development of cutaneous hypersensitivity (Grappel et al., l974) which can be demonstrated by a skin test. The skin test is a classical and widely used clinical procedure for the detection of acquired cell-mediated immunity. Skin tests are done by injecting crude or purified extract of one or more dermatophyte species intradermally. Investigations of dermatophytes’ antigens have been directed to the reactivity of different antigenic preparations and their value in detecting hypersensitivity to dermatophyte infections (Mackenzie, 1983). The studies of skin tests began in l902 when Neisser published the work of his assistant, Plato, who had prepared extracts from the Trichophyton species. He called it Trichophytin. A positive reaction shows an area of inflamed, indurated skin at the site of the Trichophytin inoculation. This reaches a peak at about 24-72 hours and then subsided.
In experimental human infections, Neves (l962) demonstrated that delayed hypersensitivity was the main response in volunteers with highly inflamed dermatophytic lesions. In chronic T.rubrum infections, he could not find a high percentage of immediate-type reactions. In cattle experimentally infected with T.verrucosum, Lepper (l972) found that the development of a well-pronounced delayed hypersensitivity was associated with the ability to eliminate the infection in four of six infected calves.
The use of Trichophytin in skin tests to aid in the diagnosis and prognosis of the disease has, however, been affected by the lack of a standardised Trichophytin (Grappel et al., l974). Different methods of preparation and purification by many investigators have led to marked discrepancies in results. Cruickshank et al.(l960) and, later, Barker et al.(l962) isolated a glycopeptide from T.mentagrophytes that gave rise to immediate and delayed hypersensitivity in humans and guinea pigs. They demonstrated that the immediate reaction was associated with the carbohydrate moiety and the delayed reaction was elicited by the peptide moiety. Later, Kaaman et al. (l976) studied three different Trichophytin preparations, viz. a purified trichophytin extract prepared by extraction with ethylene glycol, a phenol extract and a commercially available Trichophytin. The three preparations were tested on guinea pigs immunised with T.mentagrophytes in Freund’s complete adjuvant. The purified Trichophytin showed significantly delayed skin reactions in all the immunised animals but in none of the controls. It proved to be superior to the other preparations tested. In a separate study, Kaaman (l978) evaluated purified and commercial Trichophytins in 114 patients with dermatophytosis and 32 controls. Delayed reactions to the purified Trichophytin occurred in 36% of the patients but in none of the controls. Immediate reactions to purified Trichophytin were seen in 26% of the chronic patients. It was concluded that most patients with active dermatophytosis showed a delayed reaction to Trichophytin while only a few showed an immediate reaction. In contrast, chronic infection was characterised by immediate reactions and few delayed reactions.
Recently, two series of experimental infections of T.mentagrophytes were made on the forearm of a male volunteer (Nakajima, 2005). The principal mechanism of defence lay in the removal of the fungi together with keratinocytes whose turnover increased because of the eczematous reaction. It was proved that the topical application of steroid ointment suppressed the immune reactions locally; not only the clinical symptoms but also the fungicidal tissue reactions were subdued.
Immunoprophylaxis of Tinea:
Tinea is a zoonotic infection, i.e. infection that can be transmitted from animals to humans.Thus animals form a reservoir for human infection. A practical outcome of the study of immune responses to ringworm is immunoprophylaxis.
It has been reported that the level of post-infection immunity of ringworm depends on the extent of the disease and the frequency of administration of the infectious agent (Rybnikár and Oborilová, 2008). Hence, development of immunity to reinfection strongly suggests that control of ringworm by prophylactic vaccination is possible.
Immunisation of animals against ringworm has been reported by early workers and was done by inoculation by means of living or killed dermatophyte mycelia which conferred resistance to subsequent infection (Kielstein and Richter, l970 a, b; and Weiss et al., l977). A vaccine prepared by extraction of T.verrucosum mycelia ground in CO2 snow gave 50% protection in young calves (Florian et al., l964). Kielstein and Richter (l970 a,b) prepared a vaccine by extraction of T.verrucosum mycelia with 0.12N HC1. Subcutaneous or intra-cutaneous inoculation with this vaccine gave relative protection in heifers. An experiment was conducted by Weiss et al., (l977) to test active immunisation of guinea pigs with T.verrucosum antigen. The antigen used was a live homogenised suspension of T.verrucosum mycelia. Two intramuscular injections of this antigen protected the animals better than immunizing them with two subcutaneous injections.
Reports from the former Soviet Union showed that a vaccine to control cattle ringworm has been developed. Preliminary experiments in rabbits and calves have been successful and have confirmed its efficacy (Sarkisov et al., l976). The vaccine was made from live attenuated conidia of T.verrucosum strain 130 and called TF-130. The lyophilised form of the vaccine is known as LTF-130 and the manufacturers recommend that it should be administered intramuscularly in two doses at an interval of 10-14 days. The vaccine is claimed to be effective and safe. Potency is retained after 12-months storage in freeze-dried form (Sarkisov et al., l976). The protection rate was noted to be as high as 98-99% among vaccinated animals. The immunity lasted for at least 4-5 years and generally it was considered to be life-long (Sarkisov, l979). The vaccine was also reported to be effective in the treatment of cattle already infected with ringworm but not effective in emaciated animals (Petrovich 1987).
The vaccine has also been tested in controlled experiments or field trials in other European countries. In Belgium, a study was conducted by vaccinating 133 calves. A number of calves were reported to have contracted the disease when they came into contact with infected animals during the period of build-up of immunity. However, the incidence of ringworm was reduced when animals were vaccinated at an early age or when older animals were given an increased dose (Spanoghe et al., l985). Young calves on three farms in Sweden were vaccinated with 2 x 5 ml doses of LTF-130. Significant protection was observed on one farm only. When a higher dose of 2 x 10 ml was used, significant prophylactic effect was obtained on the remaining two farms (Tornquist et al., l985).
A controlled trial involving ten calves in the Netherlands showed the LTF-130 vaccine to be effective (Brethouwer, l982) and in Norway the vaccine has been evaluated and used on a large scale. Data derived from the vaccination of approximately 400,000 animals over a five-year period proved its efficacy for the immunoprophylaxis of cattle ringworm caused by T.verrucosum. During this period, epizootics of ringworm caused by other dermatophytes among the vaccinated herds were noted. A few cases of vaccination failure, adverse side effects and even death were observed. The study recommended that the vaccine for therapeutic purposes in animals with ringworm be used only according to the instructions of the vaccine manufacturers (Gudding and Naess, l986). Vaccination with inactivated vaccine gave inadequate immunity in most cases. The efficacy and safety of live dermatophyte vaccines have been demonstrated in many countries and effective control of cattle ringworm has been achieved in regions implementing systematic vaccination (Gudding and Lund, 1995). Vaccination of animals against ringworm is important in fighting the disease in animals and to control the spread of infection to humans.
Conclusion:
Ringworm, or tinea, is a contagious fungal skin disease that affects humans and animals. Animals may form a reservoir for human infection. Risk groups include young children and immune-compromised patients. Both non-specific and specific immune responses act to enable the body to abort an infection. However, specific immunity has the upper hand and cell-mediated immunity plays a major role in fighting tinea. Studies on natural and experimental infection have demonstrated that primary infection provided the animals with a degree of immunity. This has led to the conclusion that prophylactic vaccination of animals is feasible. Many investigators have proved the efficacy of vaccination in controlling transmission of infection to humans.
References
Barker, S.A.,Cruickshank, N.D., Morris, J.H. and Wood, S.R. (l962) The isolation of Trichophytin glycopeptide and its structure in relation to the immediate and delayed reactions. Immunology, 5, pp.627-632.
Brethouwer, A.H. (l982). Ringworm in Cattle. Tijdschrift voor Diergeneeskunde. 107 (18), pp.681-682.
Bronson, D.M., Desai, D.R. and Barsky, S.L. (1983). An epidemic of infection with Trichophyton tonsurans revealed in a 20-year survey of fungal infections in Chicago. J Am Acad Dermatol,8, pp.322-30.
Calderon, R.A. and Hay, R.J.(l984a). Cell-mediated immunity in experimental murine dermatophytosis: I Temporal aspects of T.suppressor activity caused by Trichophyton quinckeanum. Immunology, 53, pp.457-464.
Calderon, R.A. and Hay, R.J.(l984b). Cell-mediated immunity in experimental murine dermatophytosis: II Adoptive transfer of immunity to dermatophyte infection by lymphoid cells from donors with acute or chronic infections. Immunology,53,pp. 465-472.
Cruickshank, C.N.D., Trotter, M.D. and Wood, S.R. (l960). Studies on trichophytin sensitivity. J Inves Dermatol, 35 (4), pp.219-223.
Delves P. J., Martin S. J., Burton D. R. and Roitt I. M. (2011). Roitt’s Essential Immunology. 12th edition, Blackwell Publ.
Detandt, M. and Nolard, N. (1995). Fungal contamination of the floors of swimming pools, particularly subtropical swimming paradises. Mycoses, 38, pp.509-13.
Florian, E., Nemeseri, L. and Lovas, G. (l964). Active immunization of calves against ringworm. Magy Allatorv Lapja, l9, pp.529-530.
Grappel, S.F. (l981) Immunology of Surface Fungi : Dermatophytes. In: Comprehensive Immunology (Good and Day, Series Eds.) No.8. Immunology of Human Infection, Part 1. (Nahmias and O’Reilly, Eds.), Plenum Medical Book Co., New York and London, pp.495-524.
Grappel, S.F., Bishop, C.T. and Blank, F. (l974). Immunology of dermatophytes and dermatophytosis. Bacteriological Reviews, 38 (2), pp.222-250.
Gudding, R. and Lund, A. (1995). Immunoprophylaxis of bovine dermatophytosis Can Vet J, 36(5), pp.302-6.
Gudding, R. and Naess, B. (l986). Vaccination of cattle against ringworm caused by Trichophyton verrucosum. Am J Vet Res 47(11), pp.2415-2417.
Hay, R.J. (1997). Fungal Infections. In Skin Immune System SIS. Ed. Jan D. Bos. CRC Press, Florida, pp.593- 604.
Hay, R.J., Clayton Y.M. and De Silva, N. (1996). Tinea capitis in southeast London: A new pattern of infection with public health implications. Br J Dermatol,135, pp.955-8.
Kaaman, T. (l978). The clinical significance of cutaneous reactions to trichophytin in dermatophytosis. Acta Dermatovener(Stockholm) 58, pp.l39-143.
Kaaman, T., Stedingk, L-von. and Wasserman, J.(l976). An evaluation of delayed hypersensitivity in guinea pigs to various trichophytin preparation. Acta Dermatovener (Stockholm) 56, pp.283-287.
Kielstein, P. and Richter, W. (l970a). Versuche zur Immunoprophylaxe der Rindertrichophytie. Arch Exp Vet Med, 24 (5), pp.1205-1218.
Kielstein, P. and Richter, W. (l970b). Zur Prophylaxe der Rindertrichophytie in Grossbestanden durch aktive Immunisierung. (Vorlaufige Mitteilung). Mh Vet Med, 25 (9), pp.334-337.
Lepper, A.W.D. (l972) Experimental Bovine Trichophyton verrucosum Infection: Preliminary Clinical, Immunological and Histopathological observations in primarily Infected and Reinoculated Cattle. Res Vet Sci, 13, pp.105-115.
Mackenzie, D.W.R. (l983). Serodiagnosis. In : D.H. Howard (Ed.), Fungi Pathogenic for Humans and Animals. Part B : Pathogenicity and Detection: 1, Marcel Dekker, Inc. New York and Basel, pp.189-191.
Nakajima, H. (2005). The pathophysiology and defense mechanism against superficial and subcutaneous fungal infection. Nihon Ishinkin Gakkai Zasshi, 46(1), pp.5-9.
Neisser, A. (l902). Plato’s versuche uber die Herstellung und Verwendung von “Trichophytin”. Arch Derm Syph, 60, pp.63.
Neves, H. (l962). “Trichophytin” reaction in natural and experimental dermatophytosis and tuberculosis. Sabouraudia, 1, pp.197-202.
Petrovich, S.V. (l987). Immunity and Specific Prophylaxis against Dermatophytosis. In: Proceedings, ISHAM Consultation on Prevention and Control of Dermatophytosis with special reference to Immunity and Immunization. Oslo, Norway, pp.14-16.
Piιrard, G.E., Arrese, J.E. and Piιrard-Franchimont, C. (1996). Treatment and prophylaxis of tinea infections. Drugs, 52, pp.209-24.
Rybnikár A. and Oborilová, E. (2008). Clinical assessment of postinfection, postcontact and postvaccination immunity manifestation after experimental inoculation of calves with Trichophyton verrucosum culture. Mycoses, 51(3), pp.236-42.
Sarkisov,A.Kh.,(1979). Specific Prophylaxis of cattle trichophytosis. Veterinaria, 1, pp.40-41.
Sarkisov,A.Kh.,Nikiforov,L.K.,Pritula,A.S., Petrovich,S.V. and Jilavyan,H.A.(l976). Freeze- dried vaccine LTF-130 against ringworm of cattle. B Vses Inst Eks Vet, 25, pp.7-10.
Spanoghe,L.,Kuiper,J.D. and Brethouwer,A.H. (1985). Effectiveness of ringworm vaccine LTF-130 in cattle under field conditions. Tijdschrift voor Diergeneeskunde, 110 (23), pp.1011-1014.
Tornquist,M.,Bendixen,P.H. and Pehrson,B.(1985). Vaccination against ringworm of calves in specialised beef production. Acta Veterinaria Scandinavica,26 (1), pp.21-29.
Weiss,R.,Bohm,K.H. and Taha El Sayed,M. (1977). Immunisierungsversuche an Meerschweinchen mit einem Trichophyton verrucosum Lebendantigen. Mykosen, 20 (2), pp.54-64.